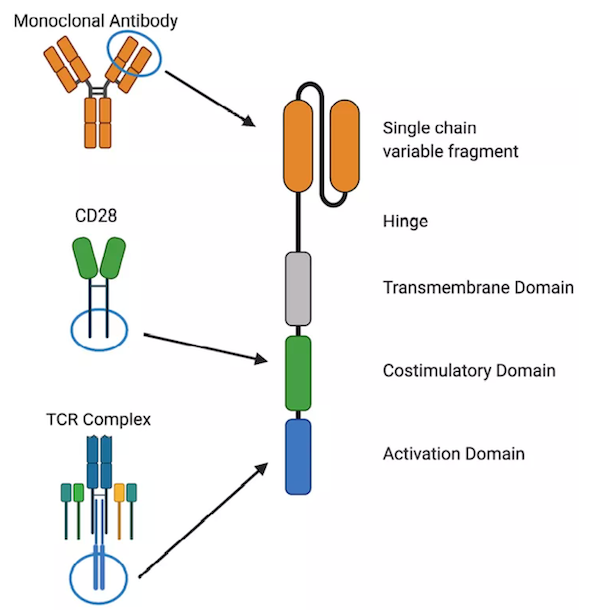
嵌合抗原受体(CARs)T细胞进行过继免疫治疗是改善癌症患者预后的一种非常有前景的方法。虽然CAR-T细胞血液系统恶性肿瘤很有效,但克服实体瘤依然困难重重,需要提高这种治疗方法的疗效。目前,有几种方法可以提高CAR-T细胞的抗肿瘤活性,包括靶向多种抗原;提高T细胞的增殖/持久性;增强肿瘤部位的归巢以及使CAR-T细胞对免疫抑制肿瘤微环境(TME)产生抵抗。通过转基因表达细胞因子或工程化的细胞因子受体来增强T细胞活化的信号已成为一种很有前途的策略,因为它不仅能提高CAR-T细胞的扩增和持久性,而且还能增强它们在免疫抑制性TME中的功能。CARs由四个组成部分组成:细胞外抗原识别结构域,最常见的是单链可变片段(scFv);结构成分,如铰链和跨膜结构域;提供维持CAR-T细胞效应器功能的共刺激信号结构域,以及CD3ζ 激活域。生理性T细胞激活需要三种不同的信号来获得效应器功能和形成免疫记忆。
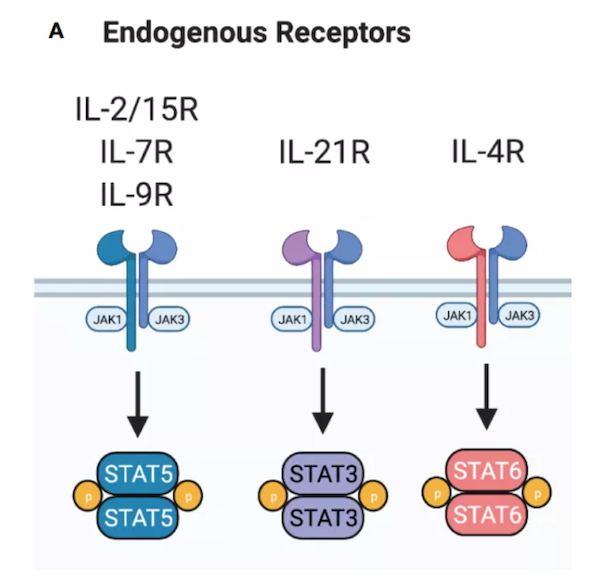
信号1(激活):T细胞受体(TCR)识别抗原后,通过CD3ζ介导的信号转导。信号2(共刺激):来自CD28或其他分子的额外信号以增强信号1。信号3由细胞因子介导,促进原始T细胞增殖分化,从而转化为效应T细胞。 第一代CAR仅通过CD3ζ提供信号1,第二代CARs也提供了信号2,通常通过CD28或4-1BB共刺激。尽管活化的CAR-T细胞能够产生细胞因子,如IL-2,但反复暴露于肿瘤细胞后,细胞因子的产生会减少。另外,一些对T细胞效应器功能很重要的细胞因子,如IL-12和IL-15,要么表达水平低,要么根本不是由T细胞产生的。由于这些局限性,研究人员开始着眼于进一步增强CAR-T细胞,通过组成性和诱导性表达细胞因子或者工程化细胞因子受体来增强信号3。细胞因子的γ链共受体家族包括IL-2,IL-4,IL-7,IL-9,IL-15和IL-21,它们在T细胞分化,增殖和内环境稳定中起着关键作用。这些细胞因子的受体包括共同的γ链(γc) 和一个各自单独的受体链,但IL-2和IL-15共享IL-2Rβ。这些细胞因子受体下游信号激活STAT家族的成员。到目前为止,IL-4和IL-9在过继性细胞治疗中的作用尚不清楚。IL-4虽然也有抗癌特性,但更多发挥的是免疫抑制作用。IL-9最初被描述为一种参与Th2应答的T细胞生长因子,虽然很少有文献评价IL-9在改善CAR-T细胞治疗中的效用,但最近发现Th9/Tc9极化的人类CAR-T细胞与传统的Th1/Tc1 CAR-T细胞相比具有更好的抗肿瘤活性。然而,关于IL-9在抗肿瘤反应中的作用存在相互矛盾的证据,需要进一步的工作来评估。IL-2和IL-15促进免疫细胞增殖和抗凋亡蛋白的转录,但自然杀伤(NK)细胞和T细胞亚群对这两种细胞因子的反应不同。高亲和力IL-2受体由调节性T细胞(Tregs)和活化的CD4、CD8+T细胞表达,而CD8+记忆T细胞和NK细胞对IL-2的反应程度较低。相反,CD8+记忆T细胞和NK细胞对IL-15的反应最为敏感,而Tregs对IL-15不反应。与单独治疗相比,第1代CD19-CAR-T细胞通过结构性表达IL-2能提高播散性淋巴瘤小鼠的无瘤生存率。然而,由于IL-2对Treg细胞的潜在扩增会阻碍抗肿瘤反应。此外,长期暴露于IL-2可促进激活诱导的细胞死亡(AICD),并且使T细胞向高细胞毒性的终末效应细胞分化,这些效应细胞虽然可有效杀死肿瘤细胞,但无法长久维持。在靶向包括CD19、GPC-3、CLL-1、GD2和IL-13Rα2的CAR-T细胞中结构性表达IL-15均显著提高了CAR-T细胞的抗肿瘤活性。IL-15对记忆T细胞亚群如干细胞记忆T细胞(Tscm)的扩增或维持可能有助于增强CAR-T细胞作用的持久性。第一代CD19-CAR-T细胞表达IL-15提高了其体内抗肿瘤活性,并允许细胞在小鼠体内存活长达110天。除了分泌性IL-15外,表达膜结合形式的IL-15是向CAR-T细胞提供激活信号的另一种方式。这种方法目前正在CD19-CAR-T细胞的1期试验中进行评估。除了CAR-T细胞外,CAR-natural killer T(NKT)细胞也被基因修饰以表达IL-15。IL15-NKT细胞维持了一个中枢记忆样细胞群,这些细胞不太容易衰竭和凋亡,从而提高抗肿瘤活性。此外,IL-15保护CAR-NKT细胞免受缺氧TME的抑制。基于这些研究,目前正在评估共表达IL-15的GD2-CAR NKT细胞在神经母细胞瘤患儿中的安全性和有效性。
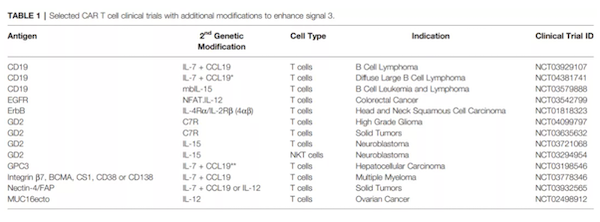
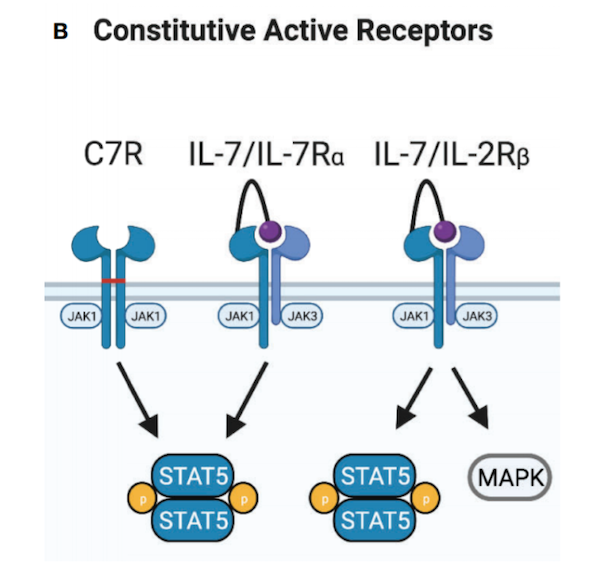
皮下注射IL-7可诱导循环中CD4和CD8+T细胞数量的剂量依赖性增加,而不会引起严重的不良事件。因此,IL-7的转基因表达已被探索作为激活CAR-T细胞的替代方法。与表达IL-2或IL-15的CAR-T细胞相比,IL-7的结构性表达提高了CD19-CAR-T细胞的抗肿瘤活性,这可能是持续的靶细胞杀伤和T细胞扩增的结果,但是这些细胞没有表现出在表达IL-15的CD19-CAR-T细胞上观察到的长期持久性。IL-7同样改善了CD20-和mesothelin-CAR T细胞,但这种作用需要C-C基序趋化因子配体19(CCL19)的共同表达。IL-7和CCL19的表达增加了CAR-T细胞、内源性T细胞和树突状细胞的肿瘤内浸润,因而具有更好的抗肿瘤活性。目前,已有多项研究进入临床阶段,以评估IL-7和CCL19表达的CAR-T细胞对淋巴瘤、多发性骨髓瘤和实体瘤的安全性和有效性。与其他γc细胞因子激活STAT5相比,IL-21更倾向于激活STAT3,同时也通过PI3K和MAPK途径介导增殖。第一代CD19-CAR-T细胞结构性表达IL-21提高了播散性淋巴瘤小鼠的整体存活率。与表达IL-2、IL-7或IL-15的CAR-T细胞相比,表达IL-21的CAR-T细胞的总存活率提高最大,尽管这些细胞表达较低水平的Bcl-2、IFN-γ, TNF-α。抗肿瘤活性的提高可能是由于维持低分化效应记忆T细胞(TEM)亚群,以及增强了长期持久性。目前,IL-21驱动的STAT3信号在CAR-T细胞免疫治疗中的疗效仍在探索中。增强T细胞活化信号3的另一种策略是通过工程化细胞因子受体的转基因表达。例如, IL-7组成性活性受体(C7R)的表达可以克服由于负反馈机制引起的IL-7Rα下调,在不需要IL-7的情况下提供持续的STAT5信号。C7R介导增强的STAT5信号以抗原依赖的方式提高GD2-和EphA2-CAR-T细胞的体外和体内抗肿瘤活性。评估表达C7R的GD2-CAR-T细胞的临床试验正在进行中。表达IL-7Rα串联IL-7是提供细胞内IL-7信号的另一种方式。此外,通过表达嵌合的IL-7Rα串联IL-17以及IL-2Rβ受体可以在CAR-T细胞中提供额外的信号,研究证明这些工程化的CAR-T细胞对TGF-β的抑制具有抗性。
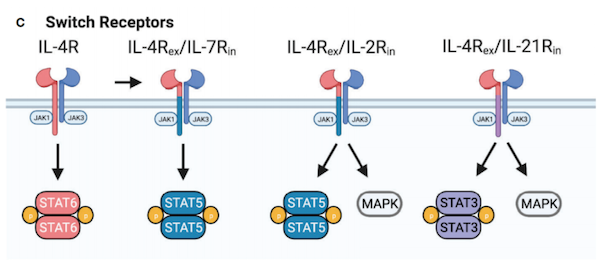
嵌合细胞因子受体或开关受体可将一种细胞因子信号转换为另一种细胞因子信号,目前正在积极探索如何劫持肿瘤或肿瘤相关细胞产生的免疫抑制细胞因子,以向CAR-T细胞提供增殖信号。结合IL-4但激活IL-7信号通路的IL-4/IL-7开关受体可以使PSCA-CAR-T细胞在体外维持其细胞溶解和增殖能力,并提高体内抗肿瘤活性。在原位乳腺癌模型中,IL-4/IL-7开关受体也提高了第二代MUC1-CAR-T细胞的抗肿瘤活性。重要的是,这些细胞能够通过IL-4依赖性的方式增殖和消除肿瘤细胞,并对远端的肿瘤再生作出反应。
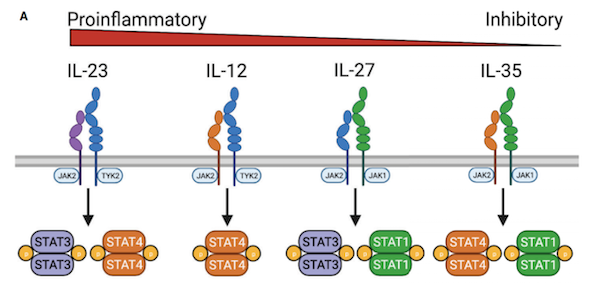
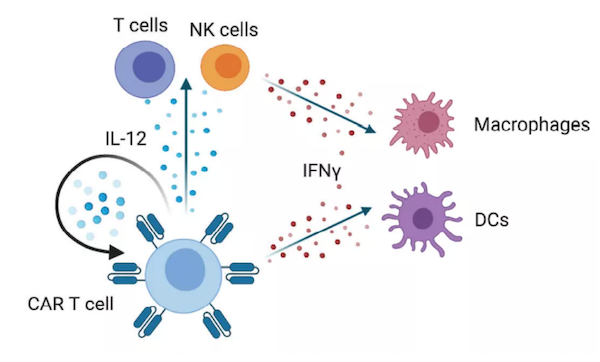
此外,在MUC1或PSMA-CAR-T细胞中表达IL-4/IL-2开关受体,通过增加IL-4存在下的STAT5和ERK磷酸化,提高其细胞溶解活性和增殖能力。目前,表达IL-4/IL-2开关受体的CAR-T细胞治疗头颈部鳞状细胞癌的1期临床试验正在进行中。IL-4/IL-21开关受体被设计成将STAT6转换成STAT3信号。这种开关受体在IL-4存在下提高了GPC3-CAR-T细胞的细胞溶解活性。很少有研究直接比较不同的细胞因子开关受体,然而,比较GPC3-CAR-T细胞中的IL-4/IL-7和IL-4/IL-21开关受体表明,在一种实体瘤模型中,IL-4/IL-21开关受体更为优越。IL-12细胞因子家族包括IL-12、IL-23、IL-27和IL-35,它们在先天性和适应性免疫应答中具有不同的作用,其中IL-12和IL-23具有促炎作用,IL-27具有促炎和抗炎作用,IL-35具有抗炎作用。由于IL-27和IL-35的主要抗炎作用,在过继性细胞治疗中,只有IL-12和IL-23有所研究。IL-12作为单药,在临床前模型中,系统给药产生了强大的抗肿瘤活性。然而,重组人IL-12的临床试验疗效有限,并且收到毒性的限制。尽管如此,表达IL-12的CAR-T细胞的仍在积极探索中。MUC16ecto-CAR-T细胞表达IL-12增强了IFN-γ的表达和持久性,提高了原位异种移植模型的存活率。研究表明,CAR-T细胞通过自分泌IL-12信号增强效应器功能,与肿瘤相关免疫细胞相互作用,耗竭免疫抑制髓系细胞,阻断肿瘤的PD-L1抑制。表达IL-12的MUC16ecto-CAR-T细胞的临床研究正在进行中。
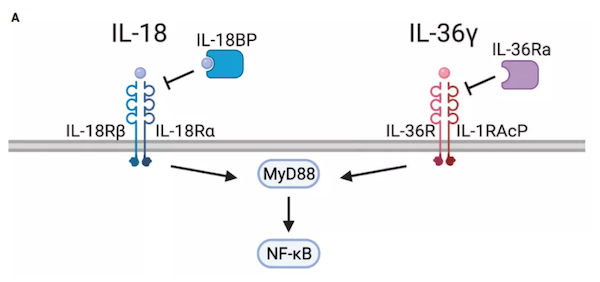
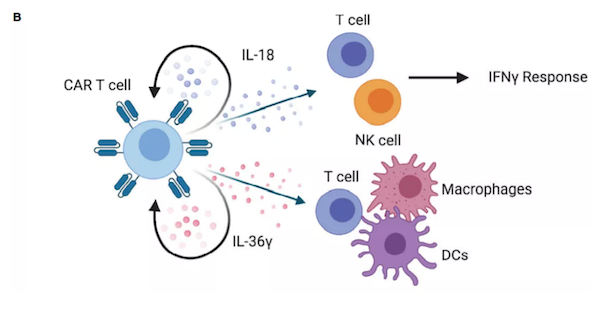
此外,工程化T细胞组成性表达IL-12可能促进T细胞功能障碍,并且在临床前模型中观察到毒性,因此设计了受活化T细胞核因子(NFAT)启动子控制的IL-12表达。NFAT启动子将IL-12表达与T细胞活化联系起来,在不降低抗肿瘤活性的情况下降低全身毒性。NFAT诱导的IL-12表达的CAR-T细胞在转移性结直肠癌的I/II期临床试验正在进行中。虽然很少有研究将IL-23与细胞治疗相结合,但最近的研究表明,这可能是改善实体瘤中CAR-T细胞功能的一种选择。IL-23诱导STAT3激活,而CAR-T细胞中的STAT3信号与慢性淋巴细胞白血病患者的响应相关。目前,STAT3信号的调节已用于改善其他临床前模型中的CAR-T细胞。IL-1超家族细胞因子包括IL-1α、IL-1β、IL-33、IL-1Ra、IL-18、IL-37、IL-36Ra、IL-36α、IL-36β、IL-36γ和IL-38,它们在先天性和适应性免疫中起着重要作用。但在临床前和临床试验中,这些细胞因子中只有一部分被认为为能够诱导抗肿瘤免疫反应。IL-18是由巨噬细胞、树突状细胞、上皮细胞以及其他细胞类型产生的一种促炎症细胞因子,与NK细胞和抗原相关T细胞上表达的由IL-18Rα和IL-18Rβ组成的异二聚体受体相互作用。IL-18通过MyD88和NF-kB发出信号,并已被证明具有促肿瘤功能,如促进血管生成、转移和增殖,但由于其在Th1反应中的协同作用,被认为在更大程度上具有抗肿瘤活性。将人类和小鼠的CAR-T细胞中表达IL-18,可以改善CAR/TCR介导的增殖、细胞因子的产生和抗肿瘤活性。一项研究表明在CD4-CAR-T细胞中表达IL-18促进了CD8-CAR-T细胞的扩增。
IL-36α, IL-36β, 和IL-36γ 是新发现的IL-1超家族成员,在临床前模型中显示出强大的抗肿瘤活性。这些细胞因子共享一个由IL-36R和IL-1RAcP组成的异二聚体受体,通过MyD88和NF-kB激活下游信号。研究表明,工程化CAR-T细胞表达IL-36γ 与未修饰的CAR-T细胞相比,提高了增殖和持久性,从而提高了抗肿瘤活性。研究表明,转基因细胞因子不仅能增强CAR-T细胞的抗肿瘤活性,而且能调节TME内的其他细胞,并能诱导或增强内源性肿瘤特异性免疫反应。然而,增强CAR-T细胞功能的继发性基因修饰可能加重CRS,因此改善CAR-T细胞扩增的方法必须与剂量限制性毒性的风险相权衡。到目前为止,大多数研究只探讨了单个细胞因子的组成性表达。在CAR-T细胞活化的不同阶段,开发能使CAR-T细胞表达一系列细胞因子的方法,不仅可以提高疗效,而且可以提高安全性。随着这些方法的发展,相信未来细胞因子将为CAR-T细胞克服实体瘤提供更加强大的助力。1. Engineered Cytokine Signaling to Improve CAR T CellEffector Function. Front Immunol. 2021; 12: 684642.